یکی از اساسیترین پرسشهایی که همواره مورد توجه بشر بوده این است که ما چگونه میتوانیم بهطور انتخابی توجهمان را به موضوع خاصی در محیط اطراف معطوف کنیم و همزمان با آن از محرکهای دیگر، که اغلب مورد توجه ما نیستند، چشمپوشی کنیم، و بهعبارتی پردازش آنها را مهار کنیم. آنچهکه ما میبینیم، میشنویم، احساس میکنیم و بهخاطر میسپاریم نهتنها به اطلاعات واردشده به حواس ما وابسته است، بلکه به ویژگیهای اطلاعاتی که به آنها توجه میکنیم نیز بستگی دارد. توجه انتخابی شنوایی عبارت است از روندی که شنونده بهطور هوشمندانه تحریکهای مورد توجه را گزینش و پردازش میکند. در ابتدا محققان بعد از جنگ جهانی دوم به بررسی موضوع توجه انتخابی شنوایی پرداختند. تلاش آنان بر این بود که دریابند چرا گاهی اوقات خلبانان هواپیماهای جنگنده، پیامهای قابل شنیدن از طریق هدفون را دریافت نمیکردند. روانشناسان علوم شناختی از جمله Broadbent (1958) آزمایههای شنوایی انتخابی مختلفی را برای درک و توضیح این موضوع مطرح کردند(1). با وجود آن که در سالهای گذشته بیشتر تحقیقات برای درک سازوکارهای توجه انتخابی بر حوزۀ بینایی متمرکز بودهاند، در چند دهۀ اخیر پژوهشگران گرایش چشمگیری به سمت سازوکارهای توجه شنوایی نشان دادهاند. آنان بر این باورند که اصول یکسانی (و شاید سازوکارهای نورونی مشابهی) در توجه انتخابی هر دو حس وجود دارند(4-2).
روش بررسی
در این مقاله مطالعات، مدلها، و نظریات بیش از 50 سال گذشته (از سال 1953 تا سال 2012) در حیطۀ توجه انتخابی شنوایی مرور شده است. به علاوه، بهطور خاص شواهد اخیر دربارۀ ارتباط تنگاتنگ بین توجه انتخابی و حافظۀ فعال (working memory) مورد تأکید قرار میگیرد. برای این منظور، از بانکهای اطلاعاتی PubMed، Scopus، ProQuest، Iran Medex، و Google scholar استفاده شد. برای جستجو در این بانکها از واژههای auditory attention، working memory، top-down، bottom-up، attention Theories، و auditory attention impairments استفاده شد. اطلاعات بهدست آمده از مطالعات مربوط مورد تجزیه و تحلیل قرار گرفت و تنها مقالاتی که به نظریهها و مبانی توجه، بهویژه توجه شنوایی اشاره داشتند، استخراج شد. بهعبارت دیگر، تمرکز نویسندگان بر مقالاتی قرار گرفت که در آنها به اصول و نظریههای اساسی، با تکیه بر آزمایشهای رفتاری، روانشناختی، علوم اعصاب شناختی، و شنواییشناسی اشاره شده بود. در نهایت، با محدود ساختن هدفمند مقالات و مطالعات مربوط، منابع مقاله حاضر استخراج شد.
نظریهها و مدلها
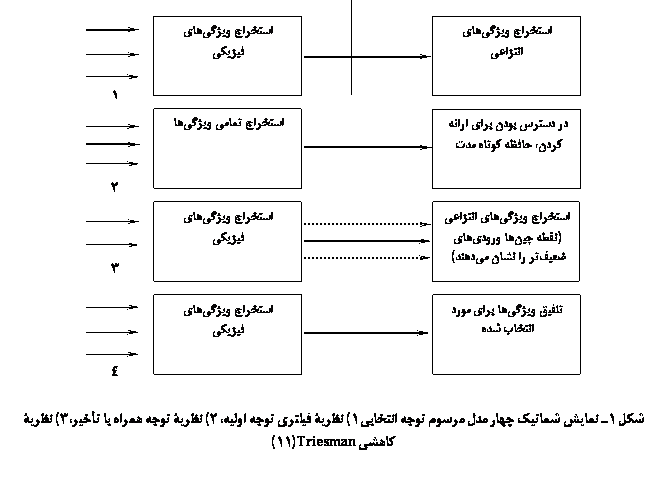
Cherry (1953 و 1954) برای اولینبار حدود 60 سال پیش در یک آزمایش شنوایی دایکوتیک نشانداد که افراد قادرند در شرایطی که بهطور همزمان پیامهای متفاوتی به دو گوش آنها ارائه شود، پیامهای هدف در یک گوش را دریافت کنند، و در عین حال محتوای پیامهای غیرهدف در گوش دیگر را مهار کنند. بعضی از این افراد تنها قادر بودند که جنس صدای گوینده در گوش غیر هدف را بهیاد آورند(5و6). Moray (1959) در تحقیق خود نشانداد که افراد حتی کلمهای را که 35 بار در گوش غیر هدف ارائه شده بود بهیاد نمیآوردند(7). نتایج این دسته مطالعات Broadbont (1958) را بر آن داشت تا مدل انتخاب اولیه (early selection model) را در توجه مطرح کند. طبق نظر وی اگر اطلاعات موجود در محیط با هدف فرد مرتبط نباشد، فیلتر خواهد شد و به سطح آگاهی (awareness) نخواهد رسید. بنابراین، فیلتر شدن براساس خصوصیات فیزیکی ظاهری از قبیل زیر و بمی، شدت و منبع صوت، در جریان گفتاری غیرتوجهی روی میدهد. از دیدگاه وی توجه انتخابی فیلتری است که مانع از پردازش اطلاعات نامربوط، فراتر ازخصوصیات فیزیکی پایهای اصوات میشود. بهنظر میرسید که وجود چنین فیلتری برای حمایت از ظرفیت محدود منابع پردازش مرکزی ضروری است تا از افزایش بیش از اندازۀ اطلاعات حسی ورودی جلوگیری شود. شکل 1 توصیف این مراحل را نشان میدهد. ولی این فرضیه که توجه انتخابی شنوایی تنها براساس خصوصیات فیزیکی محرک شنوایی باشد، مورد تردید سایر محققان قرار گرفت. در تحقیقات دیگر، شواهدی مبنی بر پردازش معنایی محرکهای ارائه شده در گوش غیر هدف مشاهده شد(8و9). پدیدۀ Cocktail Party Effect نیز شاهد دیگری بر این موضوع بود. این پدیده نشان میدهد که چطور اطلاعات غیرتوجهی میتوانند سبب برانگیختن توجه فرد شوند. تصور کنید که درحال صحبت کردن با دوستتان هستید و ناگهان نام خود را در یک مکالمۀ دیگر میشنوید. این موضوع نشان میدهد که چطور توجه شما به یک جریان غیرتوجهی فراتر از خصوصیات فیزیکی محرک برانگیخته شده است. این نتایج باعث شد تا Deutsch و Deutsch (1963) استدلال کنند که انتخاب عملاً در مراحل بالاتر پردازش اطلاعات روی میدهد. طبق مدل الهام یافتۀ نوروفیزیولوژی (neurophysiological inspired) تمام محرکهای حسی از لحاظ معنایی مورد پردازش قرار میگیرند و انتخاب (یعنی فیلتر شدن) درست قبل از آگاهشدن فرد از اطلاعات روی میدهد(10).
یکی از دلایل پایدار ماندن تناقضات دربارۀ اینکه آیا انتخاب توجهی در مراحل اولیۀ پردازش اطلاعات (جداسازی خصوصیات ابتدایی محرکها) رخ میدهد یا در مراحل بالاتر (رسیدن به سطح معنایی) ناشیاز این واقعیت است که محققان مختلف از ابزارهای متفاوتی برای بررسی آگاهی افراد استفاده میکردند.
بعضی
از محققان چند مدل شناختی و نظریههای
 واسطهای
برای شکل دادن به بدنۀ شواهد موجود در انتخاب اولیه (early) و انتخاب در مراحل
بالاتر یا با تأخیر را مطرح کردند. برای مثال Triseman و همکاران (1983)عنوان کردند فیلترهای توجهی
بهجای مسدودکردن، صرفاً پردازش اطلاعات در گوش غیرتوجهی
را کاهش میدهند (شکل 1). وی همچنین نظریۀ
بازشناسی فرهنگ لغات (dictionary
recognition units: DRUs) با
آستانههای برانگیختگی متفاوت را مطرح کرد؛ بهاین معنا که محرکهایی
مانند نام فرد یا کلمات هشداردهنده میتوانند نسبت به سایر
کلمات دیگر بهراحتی از مجرای گوش غیرتوجهی عبور
کنند(11). با اینحال شواهد
و بسترهای عصبی کافی برای توجیه بخشهای
متفاوت مدل وی ارائه نشده است. در مقابل، Johnstone و Heinz (1979) عنوان کردند عمق پردازش اطلاعات درگوش غیرتوجهی
ثابت نیست، بلکه با توجه به میزان مهارت فرد در تمایز بین
محرک هدف و غیرهدف، تغییر میکند. طبق نظریه آنها،
انتخاب میتواند در هر نقطهای از پیوستار پردازش اطلاعات
شنوایی، از مراحل اولیه تا مراحل بالاتر، رخ دهد(12). اخیراً Lavi
(2005) فرضیۀ مشابهی را دربارۀ انعطافپذیری
توجه انتخابی مطرح کرده است، اگرچه سازوکار مورد نظر وی کاملاً
متفاوت است(13).
واسطهای
برای شکل دادن به بدنۀ شواهد موجود در انتخاب اولیه (early) و انتخاب در مراحل
بالاتر یا با تأخیر را مطرح کردند. برای مثال Triseman و همکاران (1983)عنوان کردند فیلترهای توجهی
بهجای مسدودکردن، صرفاً پردازش اطلاعات در گوش غیرتوجهی
را کاهش میدهند (شکل 1). وی همچنین نظریۀ
بازشناسی فرهنگ لغات (dictionary
recognition units: DRUs) با
آستانههای برانگیختگی متفاوت را مطرح کرد؛ بهاین معنا که محرکهایی
مانند نام فرد یا کلمات هشداردهنده میتوانند نسبت به سایر
کلمات دیگر بهراحتی از مجرای گوش غیرتوجهی عبور
کنند(11). با اینحال شواهد
و بسترهای عصبی کافی برای توجیه بخشهای
متفاوت مدل وی ارائه نشده است. در مقابل، Johnstone و Heinz (1979) عنوان کردند عمق پردازش اطلاعات درگوش غیرتوجهی
ثابت نیست، بلکه با توجه به میزان مهارت فرد در تمایز بین
محرک هدف و غیرهدف، تغییر میکند. طبق نظریه آنها،
انتخاب میتواند در هر نقطهای از پیوستار پردازش اطلاعات
شنوایی، از مراحل اولیه تا مراحل بالاتر، رخ دهد(12). اخیراً Lavi
(2005) فرضیۀ مشابهی را دربارۀ انعطافپذیری
توجه انتخابی مطرح کرده است، اگرچه سازوکار مورد نظر وی کاملاً
متفاوت است(13).
طبق نظریۀ بار ادراکی (perceptual load) در توجه انتخابی که توسط Lavi وTsal (2005) مطرح شده است، منابع توجهی یک فرد همواره و بهطور کامل در پردازش اطلاعات حسی ورودی بهکارگرفته میشوند. بنابراین در شرایطی که تکلیف داده شده به فرد دشوار نباشد، منابع توجهی موجود برای پردازش سایر محرکها (مانند جریان شنوایی نامرتبط در مطالعات دایکوتیک) باقی خواهند ماند. Lavi بیان میکند که در شرایط بار کم (low-load) انتخاب با تأخیر مشاهده میشود، ولی اگر بار تکلیف خواسته شده از فرد افزایش یابد (تکالیف دشوار و پیچیده)، شرکتکنندگان منابع توجهی بیشتری را به آن اختصاص خواهند داد و منابع توجهی اندکی برای پردازش محرکهای بعدی در اختیار خواهد بود. در شرایط بار زیاد (high-load) انتخاب در مراحل ابتدایی پردازش اطلاعات رخ خواهد داد(14).
آنچه حائز اهمیت است، نقش ظرفیت حافظۀ فعال در نظریۀ فوق و توانایی متفاوت افراد در متمرکز کردن توجه انتخابی خود بر جریان شنوایی هدف است. Conway و همکاران (2001) نشان دادهاند که ارتباط نزدیکی بین حافظۀ فعال و تسخیر توجه در تکالیف شنیداری وجود دارد. افرادی که ظرفیت حافظۀ فعال کمتری دارند، بهسختی میتوانند محرکهای غیرمرتبط را فیلتر کرده و توجه خود را بر جریان هدف متمرکز کنند(15).
مثال قابل توجه دیگر، که نشان دهندۀ محدودیتهای چشمگیر در پردازش اطلاعات شنوایی است، از مطالعات ناشنوایی در تغییرات شنوایی (auditory change deafness) حاصل شده است. این اصطلاح در قیاس با پدیدۀ نابینایی در تغییرات (change blindness) به کار گرفته شده است(16). نتایج مطالعات مذکور نشان میدهد افراد نسبت به غالب اطلاعات موجود در موقعیتهای شنوایی پیچیده ناآگاهاند، بنابراین از تغییرات موجود در آن نیز آگاه نخواهند بود. افراد تنها از تغییر دربارۀ اطلاعاتی آگاهی مییابند که به آن توجه داشته باشند(17).
در حال حاضر چهار نظریۀ مشخص در ارتباط با توجه انتخابی شنوایی مطرح است که عبارتند از:
1) نظریۀ انتخاب اولیه (early selection): پردازش تحریکهایی که مورد توجه نیستند ممکن است قبل از تجزیه و تحلیل کامل حسی دستخوش تغییر و تعدیل قرار گیرد.
2) نظریۀ انتخاب همراه با تأخیر (late selection): تمامی تحریکها (حتی در سطح معنایی آنها) کاملاً پردازش میشوند، قبل از این که انتخاب یا گزینشی صورت گیرد. براساس این نظریه، نقش توجه به تنهایی کنترل دسترسی به هوشیاری، حافظه و پاسخ است. در ارتباط با هریک از این نظریهها، دو نوع الگو توسط Triesman و همکاران (1983) به این شرح مطرح شده است: الگوی فیلترینگ در مورد مدالیتۀ شنوایی که معمولاً شامل وضعیت شنوایی دایکوتیک است و در آن افراد به ورودی رسیده به گوش مورد هدف توجه میکنند و از ورودی ارائه شده به گوش دیگر چشمپوشی میکنند. دستۀ دیگر مدلهای توجه، شامل نظریههای مخزن یا ظرفیت توجه بوده و در ارتباط با توجه تقسیم شده مطرح میشوند(11).
3) نظریۀ بهرۀ توجه (attentional gain): براساس این نظریه، توجه انتخابی بهصورت یک سازوکار فیلترینگ یا بهره عمل میکند و میتواند تحریکهایی را که در مرحلۀ اول تجزیه و تحلیل حسی مورد توجه قرار نگرفتهاند مهار یا دریافت کند.
4) مدل ردیابی توجه (attentional trace): در این مدل اعتقاد بر آن است که پردازش اولیه در قشر ارتباطی شنوایی انجام گرفته و پردازش همراه با تأخیر در مناطق پیشانی مغز انجام میگیرد.
در مجموع میتوان گفت که در مورد سازوکارهای نوروفیزیولوژیک توجه انتخابی شنوایی در انسان تصور کلی بر آن است که یک سازوکار فیلترکنندۀ تطبیقی برای توجه انتخابی شنوایی وجود دارد که میتواند بهطور انعطافپذیر و پویا، بسته به میزان نیاز توجه، تنظیم شود. توجه انتخابی شنوایی به توانایی ذهنی فرد در حذف تحریکهای مداخلهکننده و انتخاب اطلاعات مرتبط صوتی بستگی دارد (Cocktail Party Effect).
مراکز دخیل در توجه شنوایی دستگاه عصبی مرکزی
برای کنترل و حفظ ردیابی توجه، این پردازش همراه با تأخیر در لوب پیشانی انجام میگیرد. گفته میشود که اطلاعات مرتبط با بخشهای تأخیری توجه، در ساختارهای عمقی قشر پیشانی شکل میگیرند. باید این نکته را در نظر داشت که در فرایند توجه، سازوکار فیلتری دستگاه محیطی شنوایی نیز دخیل است. براساس سازوکار فیلترینگ توجه، پردازش شنوایی در مکانهای مختلف از جمله در سطح دستگاه شنوایی محیطی صورت میگیرد. بسیاری از تحقیقات نشاندهندۀ اثر محیطی توجه در پتانسیلهای برانگیختۀ شنوایی ساقۀ مغز هستند. براساس این تحقیقات، حلزون مرکز پردازشهای فعال حسی بوده و این پردازشها مستقیماً با دستگاه شنوایی وابران مرتبط هستند. یافتههای بهدست آمده از نتایج گسیلهای برانگیختۀ صوتی گوش نشان دادهاند که پاسخهای واضحتر در فرکانسهایی بهدست میآیند که توجه بهآن معطوف میشود و پاسخها با دور شدن از فرکانس مورد توجه کمرنگتر میشوند. در کل، مشخص شده است که ورودیهای وابران قادر به کنترل و تعدیل سازوکار حلزونی در قالب یک روش ویژۀ فرکانسی هستند.
در بارۀ ساختارهای فعال مغز طی توجه شنوایی تحقیقاتی انجام شده است. شواهد موجود برای اثرات توجه در قشر شنوایی با استفاده از fMRI و PET بهدست آمده است. بررسیهای دیگر که در شرایط شنوایی انتخابی دایکوتیک کلاسیک انجام گرفته است، نشاندهندۀ افزایش فعالیت در شکنج گیجگاهی فوقانی قشر شنوایی است. این اثر در مجموع در قشر مقابل بهجهت توجه، بیشتر بوده و بهعنوان کوک انتخابی قشرهای شنوایی چپ یا راست، منطبق با جهت توجه، تفسیر میشود(18).
در بررسی دیگر، نتایج متفاوتی بهدست آمده است. Frith و Friston (1996) هیچگونه اثر توجه شنوایی در قشر شنوایی نیافتند، بلکه اثرات توجه بهطور آشکار در قسمت میانی تالاموس راست مشاهده شد. ساختارهای مغزی دیگر که طی وظایف توجه شنوایی فعالیت داشتند عبارت بودند از نواحی متعدد قشر پیشانی، قشرهای پیش مرکزی دوطرفه و پس مرکزی چپ، و منطقۀ حرکتی مکمل(19). تمامی پژوهشهایی که اشاره شد در ارتباط با توجه انتخابی شنوایی مطرح شدهاند. در کنار این جنبه از توجه شنوایی، جنبۀ دیگری با عنوان توجه تقسیم شده شنوایی مطرح است که در تمامی مطالعات شنوایی دایکوتیک باید به آن توجه کرد. در این قسمت سعی شده است تا به برخی از مطالعات انجام شده در شرایط شنوایی دایکوتیک اشاره شود. در مطالعهای، Hugdahl و همکاران (2000) با استفاده از PET به بررسی اثرات توجه بر وضعیت شنوایی دایکوتیک پرداختند. هدف از انجام این مطالعه نمایان ساختن تأثیر توجه بر فعالیت مغزی در شرایط شنوایی دایکوتیک بود. برای این منظور، از دو نوع محرک متفاوت استفاده شد که بهطور همزمان ارائه میشدند (هر کدام از تحریکها به یکی از گوشها ارائه میشد). محرکهای مورد نظر مشتمل بر فهرستی از هجاهای همخوانـواکه و تحریکهای موسیقی کوتاه بودند. تحریکهای گفتاری سبب فعالیت مناطق زبانی Broca و Vernike شده و تحریکات موسیقی منجر به فعال شدن قشر ارتباطی بینایی، مخچه، و هیپوکامپ شدند. یافتههای بهدست آمده بیانگر تأثیر تسهیلی توجه بر پردازش شنوایی و فعال شدن لوب آهیانهای در قالب شبکۀ توجهی بودند(20). در یکی دیگر از مطالعات، Juncke و همکاران (2003) با استفاده از fMRI به بررسی توجه شنوایی افراد هنجار در آزمایۀ شنوایی دایکوتیک پرداختند. نتایج این مطالعه نشاندهندۀ فعالیت شبکۀ عصبی توزیع یافته متشکل از پردازش اطلاعات شنوایی مسیرهای پشتی و شکمی بود. بهعلاوه، یافتهها بر تعدیل فعالیت planum temporal تأکید داشت(21). در پژوهش Talebi و همکاران (2007) روی بیماران مبتلا به سکتۀ مغزی مشخص شد که توجه تقسیم شدۀ شنوایی بیشتر از نوع انتخابی متأثر میشود و سازوکارهای کنترلکنندۀ آنها متفاوت از هم هستند(22). در توجه تقسیم شدۀ شنوایی هر دو نیمکرۀ راست و چپ فعال بوده و مناطق قشری، همچون لوبهای آهیانهای و پیشانی، و مناطق ارتباطی قشری مانند منطقۀ پیشمرکزی دوطرفه و منطقۀ حرکتی مکمل (supplementary motor area) در شکلگیری آن سهیماند(22). همچنین، در یکی از جدیدترین تحقیقات بالینی انجام شده روی سالمندان ناراضی از سمعک (علیرغم تنظیمات سمعک آنان)، مشخص شد که در جمعیت مورد نظر، اختلالات پردازشی قابل ملاحظهای از نظر بالینی در حافظه و توجه شنوایی (توجه تقسیمشدۀ شنوایی) مشاهده میشود. نتایج این تحقیق نشانگر آن است که اختلالات مذکور احتمالاً یکی از دلایل اصلی نارضایتی سالمندان از سمعک است(23).
در فرضیات جدید مرتبط با سازوکارهای توجه انتخابی شنوایی و توجه تقسیم شدۀ شنوایی، بهوجود مسیرهای کولینرژیک اشاره میشود که از ساقۀ مغز منشأ میگیرند و به تمامی مناطق و لایههای قشری منتهی شده و عمدتاًًًًً تعدیلکنندۀ اجرای وظایف مورد نیاز برای پردازش در شرایط رقابتی هستند(24). بررسیهای آزمایشگاهی در حیوانات از این فرضیه حمایت میکنند که آسیبدیدگی مسیرهای مذکور منجر به کاهش یکپارچگی ورودیهای کولینرژیک به قشر مغز و در نتیجه اختلال در توجه تقسیمشده و نهایتاً عملکردهای شناختی میشود(24).
چهار مرحلۀ اصلی در ارتباط با توجه
درک سازوکارهای توجه برای شناسایی مبنای عصبیـزیستشناختی یک تجربۀ همراه با هوشیاری ضروری بهنظر میرسد. براساس تحقیقات انجام گرفته، مشخص شده است که چهار مرحله برای توجه ضروری است: حافظۀ فعال، کنترل حساسیت بالا به پایین یا به اصطلاح مسیر بالا به پایین (top-down sensitivity control)، انتخاب رقابتی (competitive selection)، و فیلترینگ خودکار پایین به بالا (automatic bottom-up filtering) برای تحریکهای برجسته (salient stimuli)(25). هر مرحله نقشی بسیار حیاتی در توجه بازی میکند. کنترل ارادی توجه، سه مرحلۀ اولیه را شامل میشود. این سه مرحله (حافظۀ فعال، مسیر بالا به پایین، و انتخاب رقابتی) چرخهای برگشتی (recurrent loop) تشکیل میدهند و در شرایط مختلف، ارتباطات رفت و برگشتی قابل توجهی با هم نشان میدهند.
برای برقراری رفتاری مطابق با جهان پیرامون، انسان باید اطلاعات مورد نظر و موجود در محیط را در هر لحظه دریافت و پردازش کند. این اطلاعات در حافظۀ فعال ارزیابی میشوند؛ یعنی اطلاعات واردشده به آن، با جزئیات خود تجزیه و تحلیل میشوند و تصمیمات لازم براساس آن گرفته میشود. برای انتخاب اطلاعاتی که به حافظۀ فعال میرسند، سازوکارهای توجهی نقش بارزی دارند. حافظۀ فعال شکل بسیار پویایی از حافظه است که طی چندین ثانیه عمل میکند و بهطور موقتی اطلاعات انتخاب شده را برای تجزیه و تحلیل جزئیتر ذخیره میکند. انتخاب رقابتی روندی است که نوع اطلاعاتی را که در دسترس حافظۀ فعال قرار میگیرد تعیین میکند(26). مسیر بالا به پایین روندی است که قدرت نسبی سیگنال حاصل از کانالهای متفاوت اطلاعاتی را تنظیم میکند. این کانالها برای دستیابی به حافظۀ فعال رقابت میکنند(27). فیلترهای برجسته بهطور خودکار تقویتکنندۀ پاسخ به محرکهایی هستند که از نظر زمانی و فضایی یکساناند و یا از اهمیت زیستی برخوردارند. نورونها و روابط تحریکی و مهاری میان آنها تشکیلدهندۀ فیلترهای مذکور هستند(28). نتیجۀ عملکرد این چهار مرحله، افزایش حساسیت رفتاری و کاهش نهفتگی پاسخ و در نهایت، افزایش پردازش همراه با شناخت است(25). چارچوب فرض شده برای توجه در شکل 2 نشان داده شده است. این مدل براساس تحقیقات Desimone و Duncan (1995) و Miller و Cohen (2001) طراحی شده است(26و29). دستگاه عصبی مرکزی شامل اطلاعاتی در مورد جهان پیرامون و حالت درونی انسان است. در هر لحظه از زمان، اطلاعاتی که قرار است به حافظۀ فعال برسند براساس رویکرد رقابتی و بر مبنای قدرت نسبی سیگنال انتخاب میشوند. قدرت سیگنال نشاندهندۀ اثرات ترکیبیافتۀ کیفیت اطلاعات رمزگذاری شده، سیگنالهای بالا به پایین، و فیلترهای برجستۀ پایین به بالا هستند. اطلاعاتی که از بیشترین قدرت برخوردار هستند، وارد مدار حافظۀ فعال میشوند و برای کنترل این حافظه با اطلاعات موجود رقابت میکنند. اطلاعات موجود در حافظۀ فعال از طریق مسیر بالا به پایین به تعدیل قدرت سیگنال در نمودهای صعودی مرتبط میپردازند. این وضعیت، تشکیلدهندۀ یک چرخۀ برگشتی مرتبط با توجه ارادی است.
حافظۀ فعال
حافظۀ
فعال عبارت است از توانایی نگهداری «فعالانه» اطلاعات در ذهن برای انجام
فعالیتهای پیچیده از قبیل استدلال، درک مطلب و
یادگیری. این حافظه از اطلاعات محدودی طی
چندین ثانیه برخوردار است(30). این اطلاعات به
شیوهای کاملاً انعطافپذیر و بر مبنای وضعیت
درونی فرد ارزیابی شده و مورد استفاده قرار
میگیرند. در حافظۀ فعال، روند رقابتی مختلفی وجود

